Introduction
The chronic infection of hepatitis B virus (HBV), a global public health problem, is one cause of the occurrence and development of hepatocellular carcinoma (HCC) [1]. HBV genome has four overlapping open reading frames (ORFs) called C, S, P, and X. The C ORF has two genes that encode HBV core protein (HBc) and HBe protein. S ORF contains preS1, preS2, and HBs domains and responsible for the expression of three viral envelope proteins, including LS, MS, and S. P and X ORFs have viral polymerase (HBp) and HBx genes, which encode the Pol protein and viral transactivator HBx protein [2]. Among these viral proteins, HBc protein is a component of viral nucleocapsid, which not only facilitates virus replication but also contributes to the pathogenesis mediated by HBV [3]. Furthermore, increasing evidence demonstrates that, via regulating the expression and function of a variety of cellular factors, HBc also has a vital effect on the hepatocarcinogenesis during HBV infection [4–6]. For instance, HBc facilitates the growth of hepatoma cells via hTERT and Src/phosphoinositide 3-kinase (PI3K)/Akt pathways [4,5]. The viral protein is capable of enhancing the metastasis of hepatoma cells through enhancing miR-382-5p level and reducing DLC-1 expression [6]. Considering the potential importance of HBc in the development of HCC during HBV infection, further exploring the molecular mechanisms related to the dysfunction of hepatoma cells mediated by HBc may help us find novel therapeutic strategies for HBV-related HCC.
C5α receptor 1 (C5AR1) is an important component of complement systems. After binding to complement C5α, the activation of C5AR1 could regulate the differentiation and function of various target cells and participate in multiple innate immune responses [7,8]. Until now, collective evidence has proved that C5AR1 is involved in the pathogenesis of a variety of human tumors [9–11]. Via binding to C5α, which is from cancer cells or tumor microenvironment [12,13], C5AR1 could promote the malignant development of tumors with different types. Especially, Hu et al. [14] found that C5AR1 expression was significantly enhanced in HCC tissues, and it could promote the invasion of hepatoma cells via epithelial-mesenchymal transition mediated by ERK1/2. However, whether C5AR1 was involved in the development of HBV-related HCC is poorly understood.
In the present study, we explored whether HBV and virus-encoded proteins could regulate the expression of C5AR1 in hepatoma cells, and the results showed that HBc protein contributed to the upregulation of C5AR1 mediated by the virus in hepatoma cells. Besides, the molecular mechanisms related to the increased expression of C5AR1 mediated by HBc were investigated. The biological functions and associated molecules that could be affected by HBc via C5AR1 in hepatoma cells was also assessed. The relevant findings could help us better understand the role and associated mechanisms of hepatocarcinogenesis mediated by HBc during HBV infection.
Materials and Methods
1. Reagents and plasmids
The antibodies against C5AR1 were from Santa Cruz Biotechnology (Santa Cruz, CA). The antibodies against p65, phosphorylated p65, JNK, phosphorylated JNK, ERK, phosphorylated ERK, p38, phosphorylated p38, AKT, phosphorylated AKT, tumor necrosis factor α (TNF-α), interleukin (IL)-6, and IL-1β, were purchased from Bioss (Bejing, China). The antibody against Flag Tag was obtained from Proteintech (Wuhan, China). The human IL-1β enzyme-linked immunosorbent assay (ELISA) kit and Sp1 antibody were from ABclonal (Wuhan, China). The antibody against HBc was from Abcam (Cambridge, MA). Recombinant human C5α was obtained from Sino Biological (Bejing, China). The human C5α ELISA kit, TNF-α ELISA kit, and IL-6 ELISA kit were from Cusabio Biotech (Wuhan, China). Sp1 inhibitor mithramycin A was from MCE (Shanghai, China). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Santa Cruz Biotechnology), anti-goat IgG–horseradish peroxidase (HRP) (ZSJQ-Bio, Beijing, China), anti-rabbit IgG-HRP (ZSJQ-Bio), and anti-mouse IgG-HRP antibodies (ZSJQ-Bio), the nuclear factor κB (NF-κB) inhibitor BAY11-7082 (Sigma-Aldrich, St. Louis, MO), Trizol reagent (Invitrogen, Carlsbad, CA), TIANScript RT Kit (TIANGEN Biotech, Beijing, China), Matrigel solution (BD Biosciences, San Diego, CA), G418 (Promega, Madison, WI), Cell Counting Kit-8 (CCK-8) kit (Dojindo Laboratories, Kumamoto, Japan), Clarity ECL Western Blot substrate (Bio-Rad, Richmond, CA), LipoMax DNA transfection reagent (Sudgen Biotechnology, Nanjing, Jiangsu, China), and the dual luciferase reporter assay system (Promega) were obtained as mentioned previously [15–17].
HBV plasmids (pUC18-HBV1.2), the plasmids containing different HBV genes, including HBc, HBx, HBs, preS1, preS2, HBe, HBp, PGL3-Basic plasmid, and pRL-TK plasmid, were obtained as mentioned [16,17]. The plasmid containing short hairpin RNA (shRNA) targeting C5AR1 (ACTTCTAACACGCTGCGTGTT), pGPU6/GFP/Neo-C5AR1, shRNA against p65 (GGAGATGAAGACTTCTCCTCC), pGPU6/GFP/Neo-p65, and a control plasmid were from GenePharma (Suzhou, China). The gene of HBc was amplified by polymerase chain reaction (PCR) and cloned into pcDNA3.1-Flag vector to construct Flag-HBc plasmid. The primer sequences used to construct the Flag-HBc plasmid were as follows: CTAGTCTAGAATGGACATTGACCCGT ATAAAG and GGAATTCACATTGAGATTCCCGAGATTGAG.
2. Real-time PCR
The real-time PCR was performed as mentioned previously [18]. Briefly, dependent on Trizol reagent, cellular RNA was extracted from the target cells. Reverse transcription was performed using TIANScript RT kit. The primer sequences and PCR amplification conditions for GAPDH were as previously described [18]. The primers for different complement receptors were presented in Table 1. The primers for C5α were ACAGTCATAGAGTCTACAGGTGG and CCAACTGGTCAAGCGA ATCTT. The conditions for amplification were: 4 minutes at 95°C followed by 15 seconds at 95°C, 15 seconds at 60°C, and 30 seconds at 72°C for 30 cycles. The relative expression of target genes was normalized against that of GAPDH.
4. Clinical samples
The HBV-negative adjacent tissues (n=40), HBV-positive adjacent tissues (n=50), HBV-negative HCC tissues (n=50), and HBV-related HCC tissues (n=65) were collected from the Department of Pathology, the Affiliated Hospital of Xuzhou Medical University, or obtained from Shanghai Outdo Biotech Co., Ltd. The serums of health controls (HC, n=15), chronic hepatitis B patients (CHB, n=18), HCC patients with HBV infection (HBV-HCC, n=18), and HBV-negative HCC patients (non-HBV-HCC, n=20) were collected from the Department of Laboratory Medicine, Affiliated Hospital of Xuzhou Medical University.
5. Construction of the plasmids containing different regions of C5AR1 promoter
Dependent on the specific primers, five different 5′-flanking regions (−2,000/+100, −1,293/+100, −919/+100, −455/+100, and −233/+100) of C5AR1 were amplified and inserted into PGL3-Basic vector and called as PGL3-P (−2,000/+100), PGL3-P (−1,293/+100), PGL3-P (−919/+100), PGL3-P (−455/+100), and PGL3-P (−233/+100). The forward primers for the C5AR1 promoter with different regions were as follows: CATGTTACCGCCTGATCTGTCAG, CATGATTGAGTCTTGGTGTTGTC, CAT GGATTATAGGTGTGAGCCATTGC, CATGCAAAAAATTAGCCGGGGGTG, CAT GGCCGAGCCTGCACAGGAGC, respectively, the reverse primer was CATGAG AAGGACCCACTGGAG. The conditions for amplification were as follows: 4 minutes at 95°C followed by 30 seconds at 95°C, 30 seconds at 60°C, and 45 seconds at 72°C for 40 cycles. The PGL3-P (−1,398/+100) mutants, which carried one substitution (ACCAGAGTGAT) in the potential binding site of NF-κB (AGGAGCTTCCT) in the C5AR1 promoter region, were constructed with the overlapping extension PCR.
6. Luciferase reporter gene assays
After cultured for 24 hours, the target cells were co-transfected with the PGL3 plasmids containing different regions of C5AR1 promoter along with pRL-TK plasmid. Then, the cells were collected and lysed in passive lysis buffer, after transfection with different plasmids for 24 hours. The results were obtained using the dual luciferase reporter assay system as followed by the manufacturer’s instruction.
7. Western blot analysis
Total proteins in the target cells were extracted and subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Next, the target proteins were transferred onto polyvinylidene difluoride (PVDF) membranes. After blocking by 5% milk in Tris-buffered saline for 2 hours at room temperature, the PVDF membranes were incubated with different primary antibodies at 4°C overnight. After that, the PVDF membranes were incubated with HRP-conjugated secondary antibodies for 2 hours at room temperature. The bands of western blot were obtained using Clarity ECL Western Blot substrate.
8. Immunohistochemistry analysis
The immunohistochemistry (IHC) was performed as previously described [16]. The target tissues were firstly fixed with 4% formaldehyde, embedded in paraffin, and then sectioned. The tissue sections were further deparaffinized and rehydrated. After incubated with 0.01 M sodium citrate to retrieve the antigen, the tissue sections further treated with 3% H2O2, and blocked with 10% goat serum. Sequentially, the tissue sections were incubated with C5AR1 or HBc antibodies overnight. Then, HRP-conjugated antibodies were used to incubate with the tissue sections for 2 hours. Next, the tissue sections were stained by 3,3′-diaminobenzidine, and terminated with double distilled water. Finally, the tissue sections were counterstained with hematoxylin.
The relative expression of C5AR1 and HBc in target tissues was analyzed as described by Wang et al. [20]. Briefly, to evaluate the expression levels of C5AR1, the intensity of staining of hepatoma cells was scored as below: 0 (no staining), 1 (weak staining), 2 (moderate staining), and 3 (strong staining). An intensity score was no less than 2 was regarded as high expression, an intensity score was no more than 2 was considered as low expression.
9. Immunofluorescence analysis
The target cells were seeded on coverslips for 24 hours. Sequentially, the coverslips were fixed for 5 minutes with ice-cold acetone and blocked with 5% bovine serum albumin in phosphate-buffered saline (PBS) for 30 minutes. Then, the coverslips were incubated with p65 antibodies for 2 hours. After washed with PBS, the coverslips were incubated with Alexa Fluor 594-conjugated antibodies for 2 hours. The nucleus was stained with DAPI for 10 minutes. The results were observed using the Olympus microscopy (Olympus Optical Co Ltd, Tokyo, Japan).
10. ELISA
The concentrations of C5α in serum were detected by the human C5α ELISA kit. The concentrations of TNF-α, IL-6, and IL-1β in culture supernatants were measured by the human TNF-α, IL-6, and IL-1β ELISA kit. The manipulation was following the manufacturer’s operating manual.
11. Animal transplantation
The experiments of animal transplantation were approved by the Animal Care and Use Committee of Xuzhou Medical University, and also performed by following the ethical principles for scientific experiments on animals of the Swiss Academy of Medical Sciences. Briefly, the 4-week old female BALB/c nude mice, which were purchased from Shanghai Laboratory Animal Co., Ltd. in Shanghai, China, were fed in a specific pathogen-free and temperature-controlled condition with 12-hour light/dark cycles. After transfection with C5AR1 shRNA or control plasmids for 48 hours, HepG2-HBc cells were resuspended in PBS with a concentration of 1×107/mL. Next, 100 μL cell suspension along with 100 μL Matrigel solution was injected into the mice at the shoulder. After fed for 14 days, the mice were killed, and the tumors were excised and analyzed as previously described [19].
12. Cell viability assay
The cell viability assay was described as previously. A total of 100 μL cell suspensions with 6-parallel wells were put into 96-well plates. After cultured for 24 hours, 48 hours, 72 hours, and 96 hours, the cells were treated with CCK-8 reagent, the relative growth rate of target cells was assessed with optical density values at 450 nm.
13. Plate clone formation assay
The cells with 3-parallel wells were added into a 6-well plate. After culture for 2 weeks, the cells were washed by PBS and stained with crystal violet staining solution, and the clone formation efficiency was calculated.
14. Transwell assay
The target cells were resuspended in serum-free medium and placed in the upper chamber of the Transwell plate. The chemoattractant, 100 μL medium containing 10% fetal bovine serum, was placed at the bottom chamber. After incubated for 24 hours at 37°C, the cells that did not pass through the membrane in the upper chamber were removed. The cells, passed over the membrane, were named as the migrated cells. After fixing with 4% paraformaldehyde, the migrated cells were stained with crystal violet, and then observed using the Olympus microscopy.
15. Wound healing assay
The target cells were cultured in 12-well plates. After the cells reached 90% confluence, a wound was created by a micropipette tip. The results were detected using the Olympus microscopy, and migration distances were calculated.
16. Statistical analysis
The data used in the study were analyzed by SPSS ver. 19.0 software (IBM Corp., Armonk, NY), and presented as means±standard deviation, analyzed by t test, One-way ANOVA, or Mann-Whitney test. The relative expression of C5AR1 and HBc proteins, and the correlation of C5AR1 and HBc proteins expression in HBV-related HCC tissues detected by IHC analysis were analyzed by chi-square test. The results of semiquantitative western blot analysis were measured by ImageJ software (NIH, Bethesda, MD). The p < 0.05 was considered statistically significant.
Results
1. HBc facilitates the expression of C5AR1 in HBV-related hepatoma cells
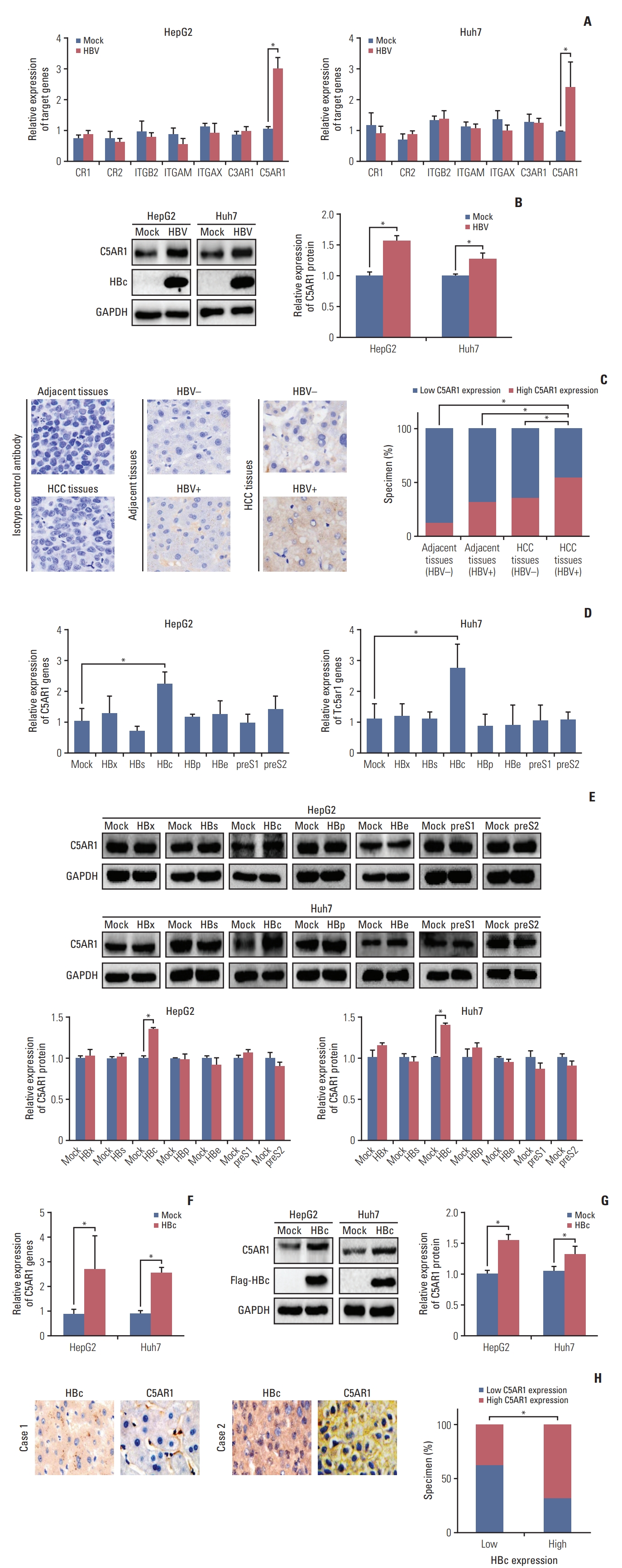
We firstly assessed whether HBV could regulate the expression of different complement receptors in hepatoma cells. HBV and control plasmids were transfected into two hepatoma cell lines HepG2 and Huh7. The mRNA expression of seven types of complement receptors, including CR1, CR2, ITGB2, ITGAM, ITGAX, C3AR, and C5AR1, were detected by real-time PCR. As the results showed in Fig. 1A, only the expression of C5AR1 mRNA was increased in HBV-positive hepatoma cells. The enhanced expression of C5AR1 protein in hepatoma cells induced by HBV was also confirmed by western blot (Fig. 1B).
The expression of C5AR1 protein in the HBV-negative adjacent tissues, HBV-positive adjacent tissues, HBV-negative tumor tissues, and HBV-positive tumor tissues was measured by IHC analysis. C5AR1 expression was observed in five out of 40 HBV-negative adjacent tissues (12.5%), 16 out of 50 HBV-positive adjacent tissues (32.0%), 18 of 50 HBV-negative tumor tissues (36.0%), and 36 of 65 HBV-positive tumor tissues (55.4%) (Fig. 1C). These results indicated that the expressions of C5AR1 in HBV-positive tumor tissues were higher than those in HBV-negative adjacent tissues, HBV-positive adjacent tissues, and HBV-negative tumor tissues.
We next measured the effect of different viral genes in C5AR1 expression. Only HBc was observed to significantly induce C5AR1 expression at mRNA and protein levels measured by real-time PCR and western blot (Fig. 1D and E). The expressions of C5AR1 in hepatoma cells stably transfected with HBc with Flag Tag (Flag-HBc) or control plasmids were also investigated. We found that, compared to control cells, C5AR1 expression was increased in HBc stable-expressing cells (Fig. 1F and G).
We also assessed the relationship between HBc and C5AR1 expression in HBV-related HCC tissues. The higher C5AR1 expression was observed in nine out of 24 HBV-positive HCC tissues with HBc low expression (37.5%). But it was found in 28 out of 41 HBV-positive HCC tissues with HBc high expression (68.3%) (Fig. 1H). Taken together, these results suggested that, HBc was associated with C5AR1 expression in HBV-related HCC tissues.
2. HBc promotes the activity of C5AR1 promoter via the NF-κB pathway in hepatoma cells
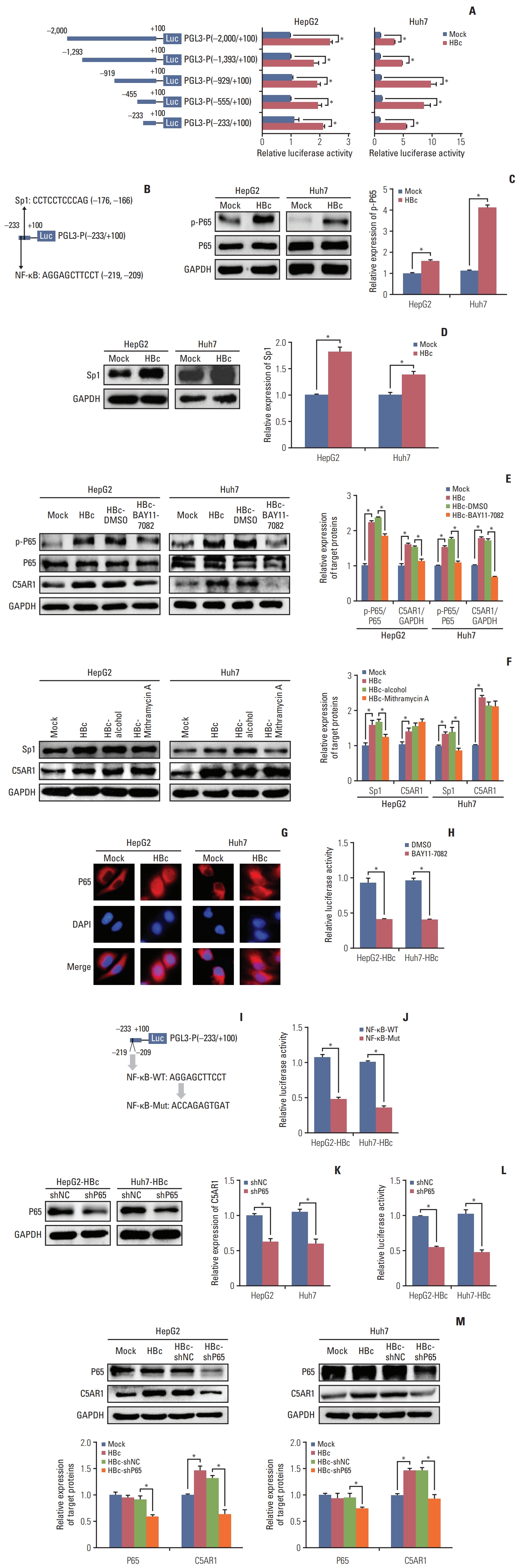
To explore the mechanisms related to C5AR1 expression mediated by HBc, five C5AR1 promoter regions, including −2,000/+100 [PGL3-P(−2,000/+100)], −1,293/+100 [PGL3-P (−1,293/+100)], −919/+100 [PGL3-P(−919/+100)], −455/+100 [PGL3-P(−455/+100)], and −233/+100 [PGL3-P(−233/+100)] were cloned into the PGL-3 vectors. These vectors were co-transfected with HBc or control plasmids into hepatoma cells for 24 hours to assess the effect of HBc on the activation of different regions of the C5AR1 promoter. As the results are shown in Fig. 2A, HBc could activate PGL3-P(−2,000/+100), PGL3-P(−1,293/+100), PGL3-P(−919/+100), PGL3-P(−455/+100), and PGL3-P(−233/+100). These results indicated that the regulatory site regulated by HBc was located in the region (−233/+100) of the C5AR1 promoter.
Next, using JASPAR database, the transcription factor binding sites in the promoter regions (−233/+100) of C5AR1 were predicted. Among the predicted transcription factors binding sites, we found that one potential binding site of NF-κB was located in the region (−214/−204) of C5AR1 promoter, and one potential binding site of the transcription factor Sp1 was located in the region (−171/−161) of C5AR1 promoter (Fig. 2B). It has been reported that HBc could transactivate the NF-κB pathway and enhance Sp1 expression to increase IL-6 expression and hepatocarcinogenesis [21], and we speculated that HBc might activate the C5AR1 promoter through NF-κB and Sp1. As the results are shown in Fig. 2C and D, we found that HBc could increase the phosphorylation of p65 protein and the expression of Sp1 in hepatoma cells. To further investigate whether HBc activates the promoter of C5AR1 via the NF-κB and Sp1, the NF-κB inhibitor BAY11-7082 and Sp1 inhibitor mithramycin A were given to HBc-positive hepatoma cells. The results showed that BAY11-7082 could significantly suppress C5AR1 expression (Fig. 2E), but mithramycin A cannot inhibit the levels of C5AR1 (Fig. 2F). These results indicated that NF-κB but not Sp1 could promote C5AR1 expression in hepatoma cells mediated by HBc.
The activation of the NF-κB pathway is related to the transfer of p65 into the cell nucleus [22]. We investigated the location of p65 in HBc-positive hepatoma cells and control cells. The results showed that, compared with control cells, and increased nuclear p65 expression was found in HBc-positive hepatoma cells (Fig. 2G). We also found that the activity of C5AR1 promoter was inhibited after the HBc-positive hepatoma cells was given with NF-κB pathway inhibitor BAY11-7082 (Fig. 2H). Moreover, we observed that the activity of PGL3-P (−233/+100) mediated by HBc was declined when the potential NF-κB binding site sequences (−214/−204) were mutated in hepatoma cells (Fig. 2I and J). The specific shRNA targeting p65 was also transfected into HBc-positive hepatoma cells (Fig. 2K), and the results showed that p65 knockdown could significantly decrease the activity of the C5AR1 promoter (Fig. 2L). Besides, after HBc-positive hepatoma cells were given with p65 shRNA, and the expression of C5AR1 protein mediated by HBc was also declined (Fig. 2M). Together, these results suggested that the expression of C5AR1 mediated by HBc relied on the NF-κB pathway in hepatoma cells.
3. C5AR1 mediated by HBc enhances the activation of signal pathways as well as the expression and secretion of IL-6 in hepatoma cells
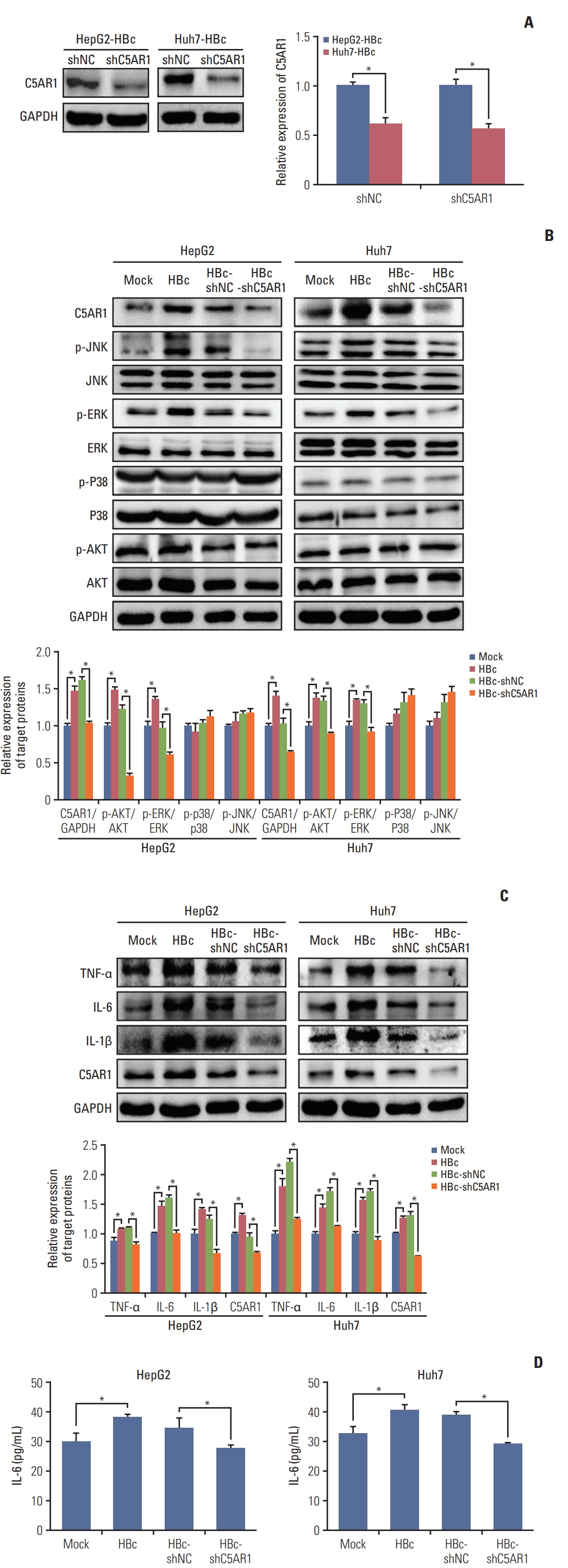
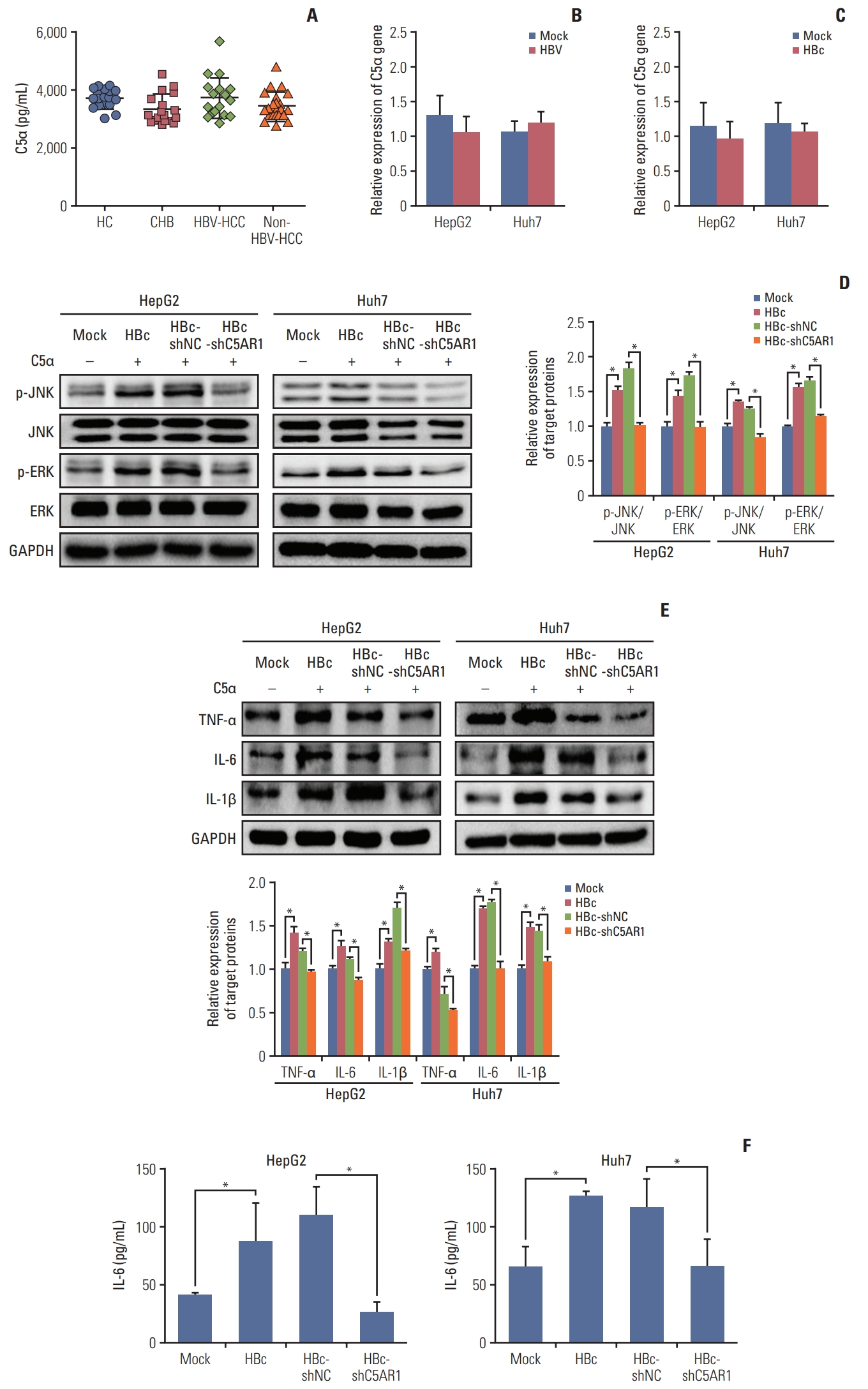
Current studies show that C5AR1 facilitates the activation of the JNK, ERK, p38, and PI3K pathways in different kinds of cells [23–25]. We next analyzed whether HBc could activate JNK, ERK, p38, and PI3K pathways via C5AR1. To better investigate the role of C5AR1 in HBc-positive hepatoma cells, the specific shRNA plasmid targeting C5AR1 was constructed, and we found that C5AR1 shRNA significantly inhibited the expression of C5AR1 protein in HBc-positive hepatoma cells (Fig. 3A). In HBc-positive hepatoma cells, the phosphorylation levels of JNK and ERK were found to be increased. Compared to control cells, no changes were found in p38 and PI3K pathways in the HBc-positive cells. After the treatment of HBc-positive cells with C5AR1 shRNA, the phosphorylation levels of JNK and ERK were significantly declined in HepG2 and Huh7 cells (Fig. 3B). Taken together, these results suggested that the activation of JNK and ERK pathways mediated by HBc mainly relied on C5AR1 in hepatoma cells.
C5AR1 facilitates the expression of inflammatory cytokines TNF-α, IL-6, and IL-1β in different cells [23,26]. Furthermore, HBc is also found could promote the gene expressions of these inflammatory cytokines in hepatoma cells [27]. We next examined whether HBc could promote the expression of TNF-α, IL-6, and IL-1β via C5AR1. As shown in Fig. 3C, when detected these inflammatory cytokines by western blot, we found that HBc could increase the expression of TNF-α, IL-6, and IL-1β in HepG2 and Huh7 cells. However, when we measured the expression of these inflammatory cytokines in culture supernatants of hepatoma cells by ELISA, only the IL-6 could be detected (Fig. 3D). After C5AR1 shRNA were given to HBc-positive hepatoma cells, the expression of IL-6 in cell culture supernatants was decreased. These results implied that, although hepatoma cells could express TNF-α and IL-1β, the expression levels of TNF-α and IL-1β is very low and this leads to the secretion of TNF-α and IL-1β from hepatoma cells cannot be detected. Except these, we found that HBc could promote the expression of TNF-α and IL-1β in hepatoma cells, but increased TNF-α and IL-1β mediated by HBc in hepatoma cells also cannot be detected by ELISA.
4. C5AR1 contributes to the growth and migration of hepatoma cells mediated by HBc
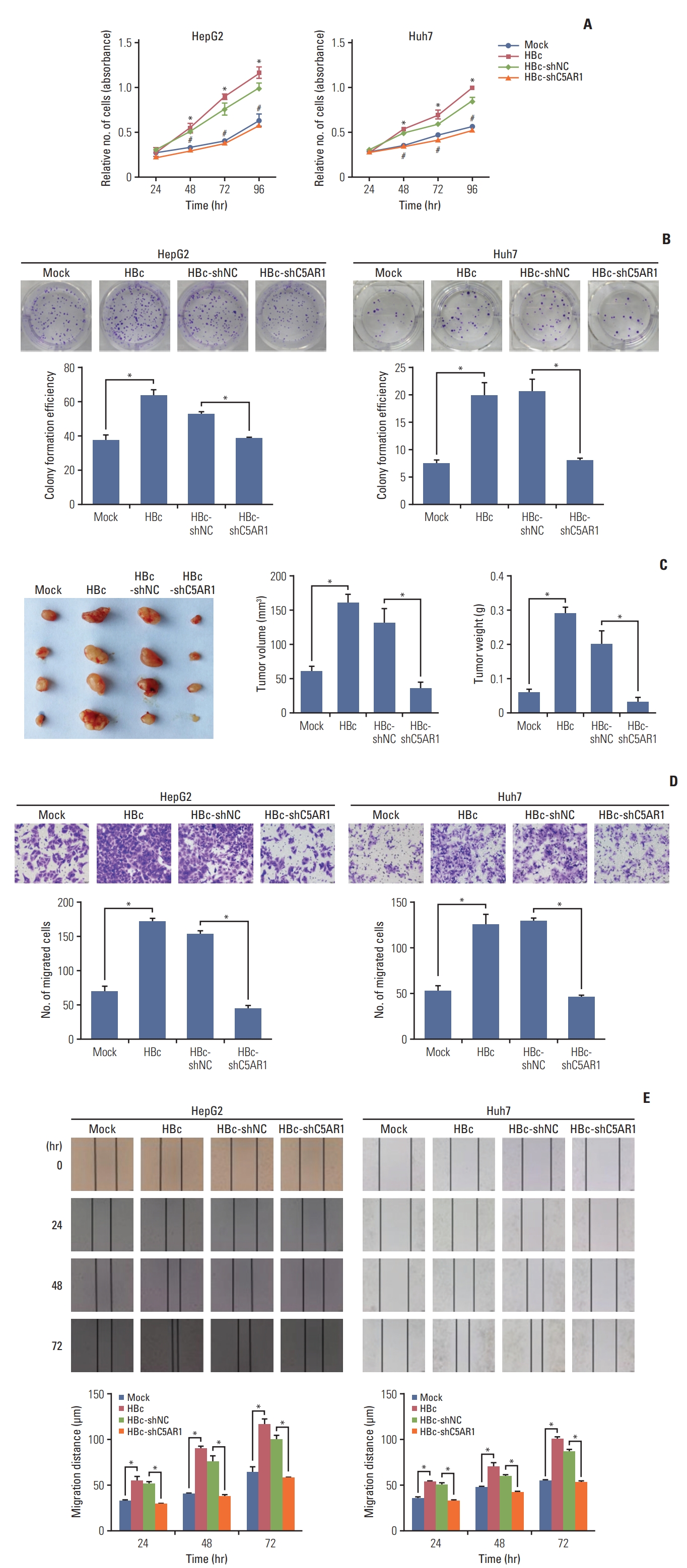
HBc has been shown to facilitate the growth and migration of the hepatoma cells [5,6]. Besides, current studies have suggested that C5AR1 played important roles in the growth and migration of different human tumor cells [9,14,25]. We examined whether C5AR1 was related to the growth and migration of hepatoma cells mediated by HBc. The results of the cell viability assay, as well as plate clonal formation assay, showed that HBc-positive hepatoma cells exhibited higher growth efficiency than control cells. After treated HBc-positive hepatoma cells with C5AR1 shRNA, the cellular growth mediated by HBc in hepatoma cells was declined (Fig. 4A and B).
We also assessed the effect of C5AR1 on the development of HBc-positive hepatoma cells in vivo. After treated HepG2-HBc cells with C5AR1 shRNA or control plasmids for 48 hours, these cells were injected into BALB/c nude mice subcutaneously. As shown in Fig. 4C, the tumor volumes, as well as weights of HepG2-HBc cells, were higher than those of control cells. After HepG2-HBc cells were treated with C5AR1 shRNA, the ability of HepG2-HBc cells to form tumors was significantly lower than that of the cells transfected with control plasmids in nude mice (Fig. 4C). Taken together, the results indicated that C5AR1 could promote the development of hepatoma cells mediated by HBc in vivo.
Sequentially, the Transwell and wound healing assay were utilized to assess the role of C5AR1 in cell migration mediated by HBc. The results showed that HBc could promote the migration of hepatoma cells. After treated HBc-positive cells with C5AR1 shRNA, the migration efficiency of hepatoma cells mediated by HBc was inhibited (Fig. 4D and E). Together, these results suggested that C5AR1 was involved in the growth as well as the migration of hepatoma cells mediated by HBc.
5. C5α facilitates the activation of signal pathways as well as the expression and secretion of IL-6 in HBc-positive hepatoma cells via C5AR1
The activation of C5AR1 is mainly based on the interaction with C5α [9,25]. We are interested in investigating whether C5α promotes the activation of signal pathways and the expression of mentioned cytokines in HBc-positive cells via C5AR1. The expression of C5α in serum was firstly assessed in HC, patients with chronic HBV infection, HBV-negative HCC patients, and HBV-positive HCC patients. However, no significant changes were found among patients in these four groups (Fig. 5A). C5α gene has been reported to be expressed in hepatoma cells [28], we were interested in detecting the expression of the C5α gene in HBV-positive cells, HBc-positive cells, and associated control cells by real-time PCR. The results showed that, compared with those in control cells, no changes in the expression of the C5α gene were found in HBV-positive cells and HBc-positive cells (Fig. 5B and C). Together, these results indicated that HBV has no significant impact on the expression of C5α in hepatoma cells.
To further determine whether C5α affects hepatoma cells via C5AR1 mediated by HBc, we added the recombinant human C5α into the medium of HBc-positive hepatoma cells. The results showed that exogenous C5α could upregulate the activation of JNK and EKR pathways (Fig. 5D), and promote the expression and secretion of IL-6, in HBc-positive hepatoma cells. Although we found that HBc could promote the expression of TNF-α and IL-1β in hepatoma cells by western blot, the secretion of TNF-α and IL-1β was not detected by ELISA. When the cells were given with C5AR1 shRNA, the increased levels of JNK and ERK phosphorylation and IL-6 expression and secretion stimulated by C5α were suppressed (Fig. 5E). Taken together, these findings suggested that C5α could stimulate intracellular pathway activation and increase the expression and secretion of IL-6 through C5AR1 in HBc-positive hepatoma cells.
6. C5α facilitate the growth and migration of HBc-positive hepatoma cells relied on the interaction with C5AR1
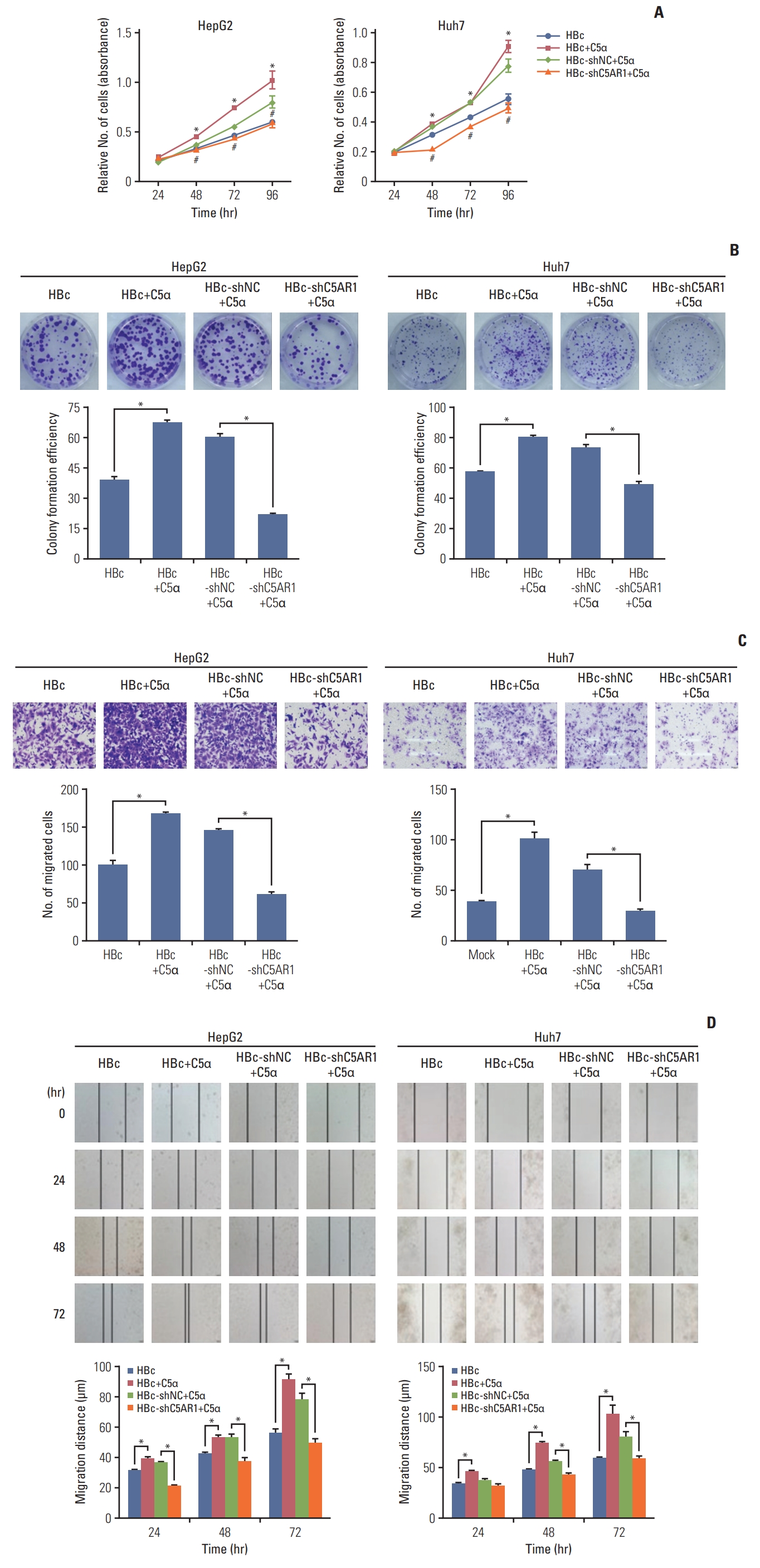
We also explored whether C5α promoted the growth and migration of HBc-positive hepatoma cells via the interaction with C5AR1. As the results showed in Fig. 6A and B, after exposure to exogenous C5α, the growth levels of HepG2-HBc and Huh-7-HBc cells were significantly increased, compared with those in cells not exposed to C5α. When the treatment of HBc-positive cells with C5AR1 shRNA, the increased growth of HBc-positive cells stimulated by C5α was declined.
Besides, when exogenous C5α was added into the medium of HBc-positive hepatoma cells, the migration of the cells significantly increased, while inhibiting C5AR1 expression of HBc-positive cells with C5AR1 shRNA, the migration of HBc-positive cells induced by C5α was inhibited (Fig. 6C and D). In summary, these results suggest that the role of C5α on proliferation and migration of HBc-positive hepatoma cells mediated mainly depends on its interaction with C5AR1.
Discussion
C5AR1 is related to the development of different human cancers [9,11,14]. However, the roles and associated molecular mechanisms of C5AR1 mediated by HBV in hepatoma cells are not fully assessed. In the present study, we discovered that HBV could promote C5AR1 expression through HBc in hepatoma cells, and the upregulation of C5AR1 mediated by HBc mainly relied on the NF-κB pathway. Moreover, C5AR1 was observed to promote the activation of intracellular signal pathways as well as the expression and secretion of IL-6, and further contribute to the growth and migration of hepatoma cells mediated by HBc.
It is known that HBV contributes to the development of HCC, but the exact mechanism of hepatocarcinogenesis mediated by the virus is still not well clear. HBc protein has the capability of forming the viral nucleocapsid and participating in HBV replication [3]. Besides, increasing evidence has demonstrated that HBc could regulate various biological processes by interacting with different cellular factors [29], and participate in the development of HCC mediated by HBV [4,5]. In the present study, we found that HBV could increase the expression of C5AR1, and the effect of HBV on C5AR1 expression relied on HBc. The aberrant expression of C5AR1 is reported in many different tumors, and it exerts vital biological functions during the development of human cancers [9–11]. Our results imply that HBc might be capable of increasing C5AR1 expression to enhance the progression of HCC.
In the study, we found that HBc could induce C5AR1 expression at the mRNA and protein levels, while the associated molecular mechanisms were not clarified. Previous studies have been reported that HBc could regulate the expression of target genes by mediating the activation of the promoters via specific transcription factors, just like CREB and c-Ets2 [4,30]. Based on these published studies, we speculated that HBc might facilitate the expression of C5AR1 via the activation of its promoters with specific transcription factors. To better understand the mechanisms related to the increase of C5AR1 mediated by HBc, different promoter regions of C5AR1 was cloned, and our results showed that HBc was able to activate C5AR1 promoter. Based on software analysis, we found that one potential NF-κB and Sp1 binding site was located in the core region of C5AR1 promoter, and the increased activation of NF-κB and expression of Sp1 mediated by HBc also be observed in hepatoma cells. Based on specific inhibitors, we found that the expression of C5AR1 mediated by HBc was mainly relied on NF-κB but not Sp1. Moreover, relied on the mutation in the potential NF-κB binding site, we found that HBc could facilitate the activation of C5AR1 promoter via the potential NF-κB binding site. Besides these, based on the inhibitor or the shRNA targeting p65, we also demonstrated that HBc could promote C5AR1 expression via the NF-κB pathway. Current studies have reported that HBc had the capability of increasing IL-6 expression through the NF-κB pathway [21]. These studies suggested that HBc might play vital roles in regulating a variety of biological functions to facilitate the development of HCC, via promoting the activation of the NF-κB pathway, to induce the expression of multiple genes, including C5AR1 and IL-6.
The activation of C5AR1 is capable of promoting the activation of JNK and ERK pathways to regulate various cellular functions in different kinds of cells [23,31]. Besides, current studies have indicated that HBc was capable of activating different signal pathways in hepatoma cells [5,21], while how HBc activates these two signal pathways is still not clear. In the study, we found that HBc could activate JNK and ERK pathways through C5AR1. The development of HCC involves the deregulation in many cellular signal pathways, and HBV is known to dysregulate multiple signal pathways to cause the progression of HCC [32]. The activation of JNK and ERK pathways induced by HBc might affect multiple biological processes to facilitate the development of HBV-related HCC.
It has been shown that C5AR1 contributes to the expression of inflammatory cytokines TNF-α, IL-6, and IL-1β in various cells [23,26]. HBc also has been shown to could induce the gene expression of different cytokines in hepatoma cells [27], while the associated molecular mechanisms are still unknown. We speculated that HBc might facilitate the expression of the cytokine via C5AR1, and the results showed that, based on C5AR1, HBc could increase the expression and secretion of IL-6 in hepatoma cells. Although TNF-α and IL-1β could be measured in hepatoma cells by western blot, the secretion of these two cytokines cannot be detected by ELISA in the target cell culture supernatants. These results implied that the resource of TNF-α and IL-1β, which could be detected by HCC patients that reported from the current published studies [33], was not from hepatoma cells, but maybe from the tumor microenvironment. Current studies indicated that the inflammation induced by cytokines is related to hepatocarcinogenesis [34]. The identified IL-6 induced by HBc through C5AR1 maybe cause hepatic inflammation and play important roles in the development of HCC during HBV infection.
Increasing evidence have indicated that C5AR1 was associated with the growth and migration of different tumor cells [9,14,25]. Besides, HBc has also been reported to be able to increase the growth and migration of hepatoma cells [5,6]. Consistent with published studies, our results indicated that HBc could increase the growth and migration of hepatoma cells. Moreover, after the treatment of HBc-positive hepatoma cells with C5AR1 shRNA, both the growth and migration of hepatoma cells were significantly decreased. These data suggested that C5AR1 plays a significant role in the growth and migration of hepatoma cells mediated by HBc.
It has been reported that C5AR1 could interact with C5α and form C5α/C5AR1 axis in various cancer cells [9,25]; In our study, we found that stimulation with exogenous C5α could facilitate the activation of JNK and ERK pathways, the expression and secretion of IL-6, and the growth and migration of HBc-positive hepatoma cells. After treated with C5AR1 shRNA, the biological function of HBc-expressing hepatoma cells stimulated by C5α could be suppressed. These results suggested that C5α could regulate the function of hepatoma cells mediated by HBc via binding to C5AR1. We found that, although HBV or HBc have no impact on the expression of C5α, the complement molecule could be expressed in hepatoma cells. Besides, C5α was reported to be expressed in various immune cells [35]. Therefore, it is reasonable to speculate that, in the hepatoma microenvironment, autocrine C5α from the cancer cells or paracrine C5α mediated by other kinds of cells, contributes to the abnormal activation of C5AR1 to facilitate the progression of hepatoma cells mediated by HBc.
In summary, our study showed that dependent on the NF-κB pathway, HBc could increase the expression of C5AR1, to increase the activation of intracellular signal pathways as well as the expression and secretion of IL-6, and facilitate the growth and migration of hepatoma cells. Our study could broaden the understanding of the biological roles and associated mechanisms related to HBc in hepatoma cells. Until now, the selective C5aR antagonists have been developed [36], and the clinical application of C5AR1-targeted drugs might be a potential therapeutic approach to control HBV-associated HCC in the future.